




Наверное, каждый из вас слышал о грозном трудноизлечимом заболевании — раке — которое может поражать многие органы у человека и животных. И возникает вопрос: могут ли растения бо-леть раковыми заболеваниями?
Болеют ли растения раком?
Рост каллуса внешне напоминает рост опухолей у животных. Но к счастью для растений, деление клеток у них всегда зависит от двух гормонов: ауксинов и цитокининов. Чтобы рост каллуса прекратился, достаточно сократить поступление хотя бы одного из них (клетки каллуса сами, как правило, не способны образовывать эти вещества). Однако, как вы помните, многие вредители и возбудители болезней растений синтезируют или ауксины, или цитокинины (в редких случаях — и те, и другие) для того, чтобы контролировать рост растительных клеток. Тогда образуются «ведьмины мётлы», галлы и прочие болезненные разрастания на теле растения. Но стоит только тем или иным способом уничтожить возбудителя, болезненный рост сразу же остановится. Таким образом, ни каллус, ни галлы не являются злокачественными раковыми образованиями.
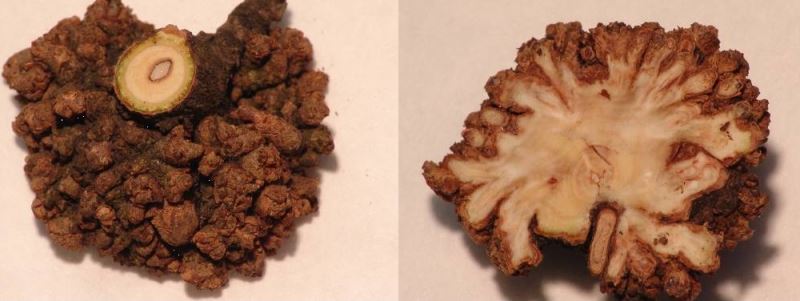
При анализе содержания гормонов в опухоли оказывается, что уровень и ауксинов, и цитокининов повышен. Каждая клетка опухоли способна самостоятельно производить эти гормоны и больше не зависит от остальных частей растительного организма.
Агробактерии — природные «генные инженеры»
Генетический материал бактерий состоит из нуклеоида (крупная кольцевая молекула ДНК, в которой хранится «основная» генетическая информация), и плазмид (более мелких кольцевых молекул ДНК с меньшей информационной ёмкостью). Свойство агробактерий поражать те или иные виды растений «запрограммировано» именно в плазмидах. В зависимости от типа заболевания эти плазмиды обозначают как Ti-плазмиды (от англ. tu-mor inducing — вызывающие опухоли) и Ri-плазмиды (от англ. root inducing — вызывающие [косматые] корни). При потере плазмид агробактерии теряют свойство вызывать соответствующие заболевания.
Плазмиды обладают целым рядом интересных и практически важных свойств. Во-первых, в одной клетке агробактерии не могут встретиться не только Ti- и Ri-плазмиды, но даже две разных Ti-плазмиды! Каким-то образом первая «поселившаяся» в бактерии плазмида не допускает проникновения и размножения других плазмид, похожих на неё.
Во-вторых, плазмиды способны передаваться от одной бактериальной клетки к другой. Как это ни парадоксально, в почве только 1 — 5% клеток свободно живущих агробактерий «вооружены» Ti- или Ri-плазмидами. Но как только начинается процесс инфекции, плазмиды активно размножаются и передаются от бактерии к бактерии.
В третьих, Ti- и Ri-плазмиды (по сравнению с другими плазмидами бактерий) имеют большие размеры: около 200 — 300 тыс. пар оснований. Это не позволяет при помощи стандартных методик отделить ДНК этих плазмид от ДНК нуклеоида, что создаёт определённые трудности в работе молекулярных биологов с плазмидами.
Какие же гены несут в себе Ti-плазмиды? Для заражения растений наиболее важным оказывается Vir-район (от англ.virulence — способность поражать [растения], патогенность), в котором закодировано довольно много генов. Постоянно работают только два гена: VirA и VirG. Белок VirA — это рецептор на особое вещество фенольной природы —ацетосирингон. Ацетосирингон выделяется при повреждении клеток растений. Белок VirA реагирует на ацетосирингон и передаёт сигнал на белок VirG, который активирует все остальные гены Vir-района. В результате: 1) клетки агробактерий плывут к месту поражения (ориентируясь по увеличению концентрации ацетосирингона); 2) Ti-плазмида начинает размножаться и передаваться другим бактериям того же вида; 3) появляются другие белки-продукты генов Vir-области (рис. 2).
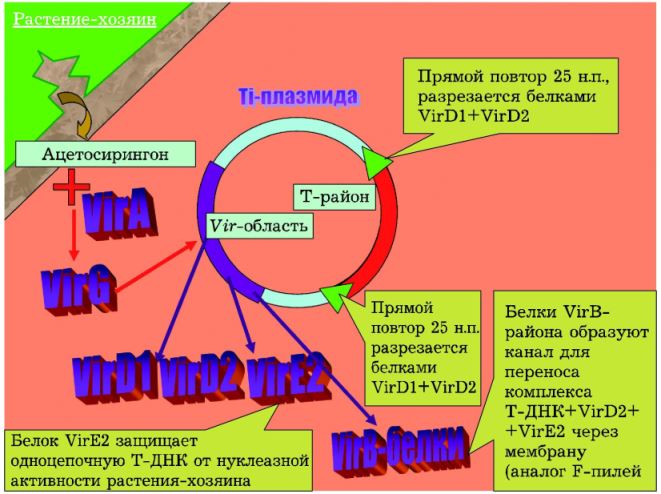
Белок VirD1 совместно с белком VirD2 находят в Ti-плазмиде определённые участки, состоящие из 25 пар нуклеотидов, и разрезают их, перебрасывая ковалентную связь с 5′-конца ДНК на белок VirD2. У Agrobacterium tumefaciens таких участков два: они ограничивают так называемый Т-район (от англ. transferred — переносящийся). Одна из цепей ДНК отделяется и уходит; таким образом, в Ti-плазмиде возникает брешь. Специальная система репарации ДНК застраивает брешь новой цепью ДНК, и из той же Ti-плазмиды можно ещё раз вырезать Т-район, Ti-плазмида в целом сохраняется.Одноцепочечная Т-ДНК, связанная с белком VirD2, в дальнейшем «одевается» при помощи белка VirE2, который не даёт ферментным системам бактериальной клетки разрушить одноцепочечную Т-ДНК.На поверхности клетки агробактерии при помощи разнообразных белков VirB образуется аппарат для переноса ДНК из одной клетки в другую. Именно белки VirB отвечают за перемещение комплекса VirD2 с одноцепочечной ДНК из клетки агробактерии в клетку растения. Белки VirE2 также перемещаются в клетку хозяина.Далее комплекс одноцепочечной Т-ДНК с белками VirD2 и VirE2 проникает в ядро растительной клетки. Белок VirD2 «надрезает» ДНК клетки-хозяина и встраивает Т-ДНК из Ti-плазмиды. Таким образом, происходит процесс встраивания чужеродной ДНК в ДНК клетки растения. После этого клетку растения можно считать генетически модифицированной. В процессе эволюции агробактерии «разработали» механизм получения генетически модифицированных клеток растения, т.е. стали природными «генными инженерами».
Что содержится в Т-районе
Гены, которые содержатся в Т-районе, в самой клетке агробактерии не работают, поскольку у них есть только эукариотические про-моторы. Два из этих генов отвечают за биосинтез ауксинов: iaaH и iaaM. Ещё один ген — iptZ — кодирует ключевой фермент синтеза изопентениладенина (одна из форм цитокининов). Таким образом, попав в геном растения, Т-ДНК вызывает синтез как ауксинов, так и цитокининов (рис. 3). При этом клетки растения-хозяина начинают неорганизованно делиться, образуя опухоль.Однако для того, чтобы деление клеток растения приносило агробактериям пользу, необходимо, чтобы они синтезировали что-либо полезное для агробактерий. Действительно, в состав Т-района входят гены биосинтеза веществ, которые образуются из аминокислот и кетосоединений. Эти вещества получили название опинов. Ни сами растения, ни другие организмы, обитающие на растениях, не могут расщепить опин. И только агробактерии способны «переваривать» тот опин, синтез которого они вызвали.Опинов достаточно много, и каждая Ti-плазмида обеспечивает синтез своего опина (нопалина, агроцинопина, витопина, куркумопина и др.). В самой Ti-плазмиде (но не в Т-районе!) есть гены, отвечающие за «переваривание» соответствующего опина. Это объясняет, почему одна Ti-плазмида, захватив клетку агробактерии, не пускает в неё другую Ti-плазмиду, отвечающую за синтез и метаболизм другого опина.После внедрения ДНК из Т-района клетки опухоли интенсивно делятся и продуцируют именно тот опин, который способна «переварить» агробактерия, вызвавшая данную инфекцию. Если в почве обитают два разных вида агробактерий, то при инфекции первая бактерия каким-то образом не пускает другую, которая питается иным опином.

На этом основан биологический метод борьбы с агробактериальным раком. Как вы знаете, есть непатогенные агробактерии. Они также «не пускают» другие виды агробактерий к корневой системе растения, через которую и происходит поражение. Если заранее обработать растение определёнными штаммами A. radiobacter, то растение не заболеет ни корончатым галлом, ни корневым раком, ни болезнью бородатого корня.Удивительно, но у некоторых агробактерий в составе плазмид есть не один, а два или даже три Т-района, каждый из которых «обрамлён» последовательностями из 25 нуклеотидов. В случае A. rhizogenes эти участки называют ТL- и TR-районами, а у A. rubi есть TA, TB и TC соответственно. Наиболее удивительно заболевание бородатого (косматого) корня. В TR-районе содержатся те же гены, что и у остальных агробактерий. Они отвечают за синтез ауксинов, цитокининов и опинов. В TL-районе содержатся гены, отвечающие за то, чтобы неактивные формы ауксинов переходили в активные. Оказывается, для успешной инфекции достаточно только TL-района! И тогда клетки опухоли активируют «запасные» формы ауксинов самого растения, а это приводит к ризогенезу, т. е. к образованию многочисленных придаточных корней на месте опухоли.Итак, в результате переноса Т-ДНК из Ti-плазмиды в клетку растения-хозяина происходит неконтролируемое образование гормонов (ауксинов и цитокининов), что приводит к злокачественному росту клеток и дальнейшему синтезу азотсодержащих веществ — опинов. К месту опухоли притекает всё больше и больше аминокислот, но они постоянно «выводятся из оборота» растения, т. к. преобразуются в новые порции опинов, которые служат источником питания для соответствующего штамма агробактерий. «Избавиться» от чужеродной ДНК растительные клетки уже не могут. Рост клеток и синтез опинов продолжаются даже в том случае, когда агробактерии по каким-либо причинам погибли.
Получение генетически модифицированных растений при помощи агробактерий
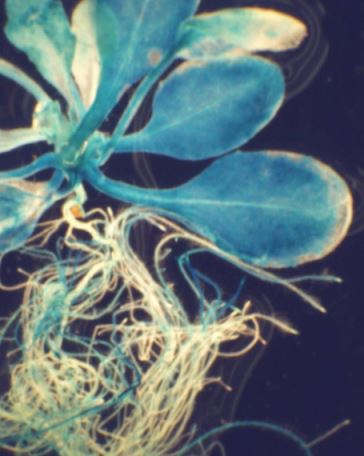
При помощи различных ухищрений с геном интереса можно получить растения, содержащие новый белковый продукт, которого раньше в клетках растений не было. Или, наоборот, можно «выключить» какой-нибудь собственный ген растения, «заставить» его работать в других органах и тканях и т.д. Это позволяет учёным детально исследовать работу генома растения. Но у генетически модифицированных растений есть и практическое применение.
